

Haplogroup G‑Z6748 is considered a rare or ‘minority’ genetic YDNA haplogroup because it descends from an already uncommon Y‑DNA series of genetic branches. It is represented today by a small, very geographically concentrated descendant cluster with a few known sub-branches and YDNA testers. Originating around 650 CE, it is predominantly associated with deep ancestral ties to Wales and neighboring parts of the British Isles and Western Europe. While primarily Welsh and British, participants who test positive for G-Z6748 are spread globally today, including in the United States, England, and across Europe.
“In Europe west of the Black Sea, Haplogroup G is found at about 5% of the population on average throughout most of the continent. The concentration of G falls below this average in Scandinavia, the westernmost former Soviet republics and Poland, as well as in Iceland and the British Isles. There are seeming pockets of unusual concentrations within Europe. In Wales, a distinctive G2a3b1 (G-P15) type (DYS388=13 and DYS594=11) dominates there and pushes the G percentage of the population higher than in England.” [1] The G-15 haplogroup is a distant ancestor of the G-Z6748 haplogroup.
“Haplogroup G-P303 (G2a2b2a, formerly G2a3b1) is a Y-chromosome haplogroup. It is a branch of haplogroup G (Y-DNA) (M201). In descending order, G-P303 is additionally a branch of G2 (P287), G2a (P15), G2a2, G2a2b, G2a2b2, and finally G2a2b2a. This haplogroup represents the majority of haplogroup G men in most areas of Europe. . . .” [2] The G-P303 haplogroup is an ancestor of the G-Z6748 haplogroup.
The following map is from an innovative study that systematically assessed the association between genetic variation in the specific regions of the Y chromosome and cardiovascular disease outcomes. While the study documented little evidence for an effect of any YDNA genetic influence on cardiovascular risk, an important secondary finding was that Y chromosome haplogroups carried by contemporary white British individuals demonstrate strong geographic structuring across Great Britain. The researchers observed that certain lineages are more prevalent in specific regions.
Illustration One: Prevalence of G2a-P15 Halpogroup by Area of Birth in Great Britain in 2022 [3]
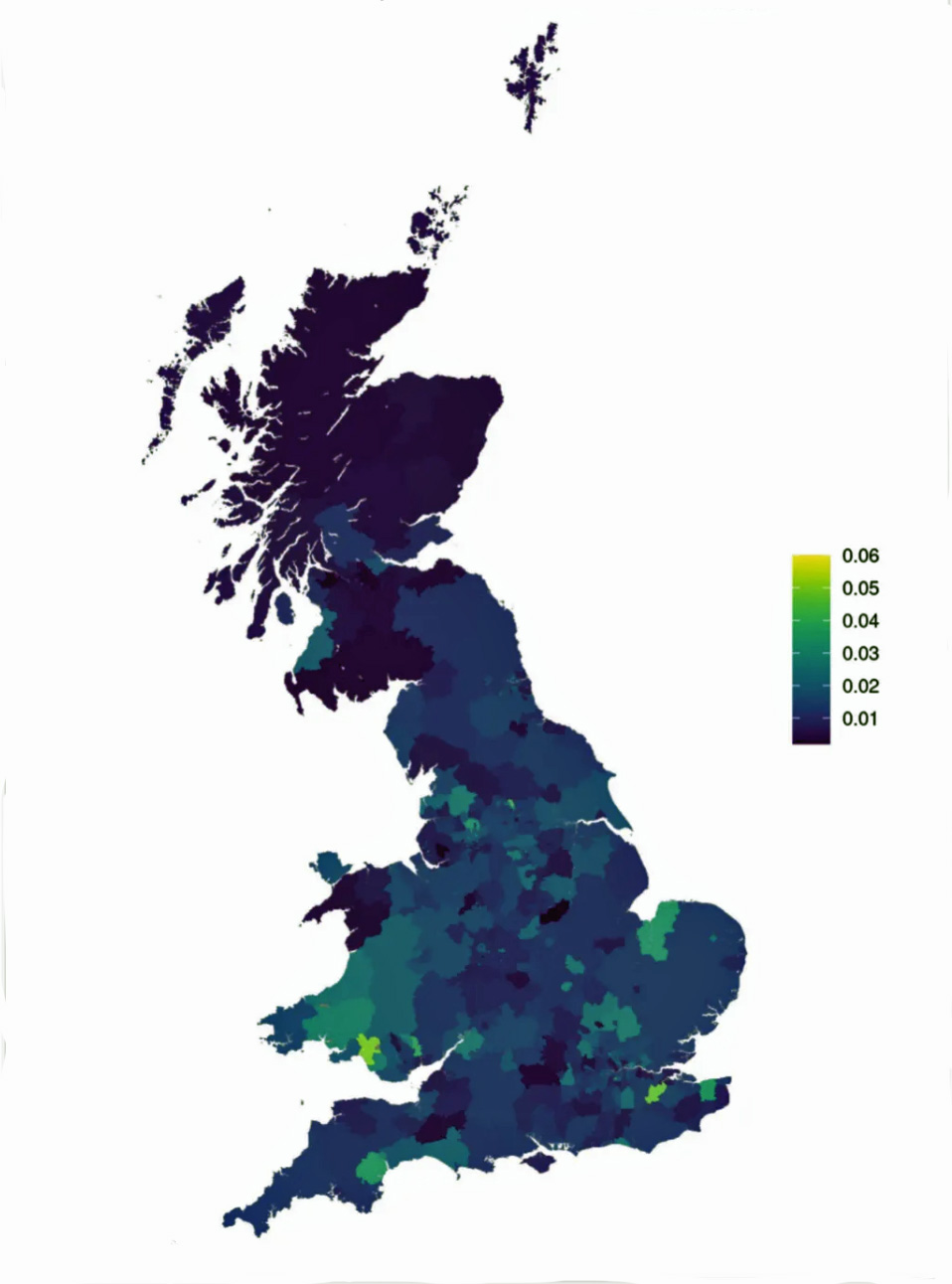
As reflected in the map, the prevalence of the G-P15 haplogroup in modern day Great Britain is rare. However, relative to other areas of the island, it is found in Wales, particularly in the central and southern areas of Wales.
This story discusses the role of a small YDNA research working group that focuses on the the genetic descendants of the most recent common ancestor of G-Z6748. Some of the results from this work group have revealed a similar patten of self reported earliest known ancestors of YDNA testers that are genetically associated wth the G-Z-6748 haplogroup.
Background Concepts
I have discussed the relationship between Single Nucleotide Polymorphisms (SNPs), Short Tandem Repeats (STRs) and haplogroups in greater detail in an earlier story. [4] Going back to basics, DNA base pairs are the fundamental building blocks of the DNA double helix, consisting of two complementary nitrogenous bases held together by hydrogen bonds. The four bases, or ‘nucleotides’, are Adenine (A), Thymine (T), Guanine (G), and Cytosine (C), eachpair specifically with antoher: A with T and G with C. These base pairs form the “complementary rungs” of the DNA ladder, dictating genetic information. Since the rungs are complimentary, one ‘rung’ is only required to define STRs and SNPs. [5]
In YDNA genealogy, SNPs, STRs, and haplogroups are three interconnected concepts that work together to trace your paternal lineage at different time depths (see table one). [6] STRs, called ‘strings’ or formally short tandem repeats, are short, 2–7 base pair sequences of DNA that repeat consecutively, known as a microsatellite. [7] SNPs, called ‘snips’ or single nucletide polymorphisms, represent a difference in a single DNA building block, or nucleotide (A, T, C, or G). [8]
Table One: ‘Strings’, ‘Snips’, and Haplogroups
“STRs are useful genealogically, to determine to whom you match within a recent timeframe, of say, the past 500 years or so, and SNPs define haplogroups which reach much further back in time. Furthermore SNPs are considered “once in a lifetime,” or maybe better stated, “once in the lifetime of mankind” type of events, known as a UEP, Unique Event Polymorphism, where STRs happen “all the time,” in every haplogroup. ” [9]
Y-DNA haplogroups are defined by the presence of a unique series of SNP genetic markers. Each branch of the genetic tree is defined by one or more specific, shared SNP mutations. Haplogroups are distinguished from one another by which of these mutations they do or do not carry. Subclades or downstream branches include the SNP mutations from prior related branches of the genetic tree but also contain one or more unique SNP mutations. [10]
Over many generations, the Y chromosome accumulates additional mutations, so haplogroups form a branching phylogenetic tree in which each branch point corresponds to a new, stable SNP event or mutation. In this sense, a haplogroup is a named genetic position on that tree. In practical terms, two men are in different haplogroup branches if they do not share the full defining SNP set for that particular branch, even though they may share older, upstream SNPs further back or upstream in the tree. In database or software contexts, haplogroups are recursive sets of groups that involve nested hierarchies. YDNA trees or phylogenetic trees list nested groups (Haplogroups) where a group is a member of another group as you go ‘down’ the subclades to more recent times. [11]
YDNA test results of individual males can be grouped into Y-DNA haplogroups based on the particular mutations found on the non‑recombining portion of the Y chromosome. These defining mutations are almost always SNPs that arose once in an ancestral male and were then passed to his male-line descendants. Table two provides an overview of the different types of SNPs referenced in a phylogenetic tree.
Table Two: Types of SNPs for a Haplogroup
| Types of SNP | Personal Example |
|---|---|
| Upstream (ancestral) SNPs | All derived SNPs that define the path from the root branch (e.g. haplogroup G-M201) down to a specific clade or terminal branch. For example, a man in G-Z6748 will also carry the defining SNPs for G-L497, and G-P303 if he is in that branch. [12] |
| Defining (haplogroup) SNPs | The specific SNP (or small set of SNPs) used as the formal label for that node. For example, the specific SNP G-Z6748 is used to name or define the G-Z6748 haplogroup branch. [13] |
| Block/cluster SNPs | On detailed trees (e.g. Big Y block trees), several SNPs may sit together at one branch because they have not yet been seen or differentiated by additional samples; all belong to that branch and are shared by everyone in the clade. For example, the G-Z6748 branch is represented by the presence of 29 SNP variants [14] |
| Terminal SNPs | The specific Y‑chromosome SNP that marks the furthest‑down (most recent) branch on the Y‑haplotree where a given man currently tests positive; it is the defining SNP of his latest known subclade. For example, my officially recognized terminal branch is G-BY211678 which is shared by 10 other YDNA testers. [15] |
| Private (novel) variants | Recently arisen SNPs seen only in one man (or one tight family cluster) so far; they are not yet used to define published haplogroups but will become defining when shared by multiple men and placed as a new twig on the tree. I have a private variant, G-FT40897, that is a new terminal branch. [16] |
FamilyTreeDNA and YDNA Work Projects
FamilyTree DNA (FTDNA) is a company that provides direct-to-consumer DNA tests for genealogy, allowing people to trace their family history through autosomal, Y-DNA, and mitochondrial DNA tests. It was founded in 2000 and is known for being one of the first companies in the field. The company offers autosomal DNA testing for broader, more recent ancestry, while Y-DNA and mtDNA tests focus on the narrower, more distant paternal and maternal lines, respectively. [17]
FTDNA is unique among major testing companies because it offers comprehensive testing options for all three types of DNA used in genealogy. The company’s strength is based on the size of their YDNA and mtDNA database and the research tools and group projects provided for genetic genealogical research. [18] Unlike many competitors, FamilyTreeDNA processes all tests in its own certified lab in Houston, allowing for potential test upgrades without requiring a new DNA sample. [19]
The three types of DNA testing provied by FTDNA are:
- Autosomal DNA (Family Finder test): This test analyzes DNA inherited from all ancestors to provide ethnicity estimates and match individuals with relatives within about five generations. Users can also upload raw autosomal DNA data from other services like AncestryDNA and 23andMe to join the matching database. [20]
- Y-DNA: Exclusively for genetic males, this series of tests traces the direct paternal line (father’s father, and so on) and can be particularly useful for surname research. Specific tests are differentiated on the number of genetic markers tested. FTDNA’s Y-DNA products have mainly differed by (1) how many Short Tandem Repeat (STR) markers they test, (2) whether they include Single Nucleotide Polymorphism (SNP)/sequence data (haplogroup resolution), and (3) how far back and how precisely they can resolve relationships and place you on the Y-tree. Historically FTDNA has offered multiple STR-only Y panels at different marker counts; currently the main marketed levels are Y-37 STR, Y-111 STR, and Big Y-700 (which also includes SNPs and STRs). [21]
- Mitochondrial DNA (mtDNA): This test traces the direct maternal line (mother’s mother, and so on) and is available for both males and females. [22]
Coupled with an extensive database for YDNA and mtDNA test results, FTDNA offers a wide variety of Y-DNA Group Projects to help further research goals of DNA testers. The group projects support genetic genealogy research, leveraging YDNA, mtDNA, and autosomal DNA results. The projects are designed to facilitate collaborative research among individuals with shared ancestry, geographic origins, or genetic interests. [23]
Joining group projects enables participants to:
- Utilize project-specific databases to break down genealogical “brick walls” and connect with distant relations;
- Compare DNA signatures and mutations within a defined subgroup; and
- Collaborate with others researching similar ancestry or geographic roots.
The group projects are associated with specific branches of the YDNA or mtDNA haplotrees, geographical areas, surnames, or other unique identifying criteria. Based on their respective area of focus, the research groups have access to and the ability to compare Y-DNA results of fellow project members to determine if they are related. These projects are run by volunteer administrators who have an interest or specialize in the haplogroup, surname, or geographical region that one may be researching.
FTDNA Group Projects
FamilyTreeDNA officially organizes its group projects into four main categories, each defined by the type of DNA analyzed and the research question it addresses (see table three) [24]
Table Three: Type of FamilyTreeDNA Group Projects
| Category | DNA Type(s) | Primary Focus | Typical Examples |
|---|---|---|---|
| Y-DNA Group Projects | Y-DNA only | Paternal-line ancestry | Surname Projects, Y-DNA Haplogroup Projects, Y-DNA Geographical Projects |
| mtDNA Group Projects | mtDNA only | Maternal-line ancestry | mtDNA Lineage Projects, mtDNA Haplogroup Projects |
| Geographical Group Projects | Y-DNA, mtDNA, and/or Family Finder | Genetic history of a specific region (country, county, city) | Finland DNA Project (largest), Greater Nordic Y-DNA Project, Brabant DNA Project |
| Family Finder Group Projects | Autosomal (Family Finder) | Descendants of a specific ancestral couple (usually 5–6 generations back) or special-interest autosomal studies | Acadian AmerIndian Ancestry Project, private family studies |
These four main project groups can alternatively be viewed in five major areas:
- Surname Projects focus on researching a specific surname, including its various spellings and branches. These usually involve Y-DNA testing because surnames are commonly passed down the paternal line, but may also include autosomal and mtDNA data when relevant.
- Haplogroup Projects target specific Y-DNA or mtDNA haplogroups or subclades. Members share a particular haplogroup and collaborate to refine its structure, migration patterns, and genetic connections.
- Geographical Projects concentrate on people from a specific region, whether by country, county, or cultural group. These may require Y-DNA, mtDNA, or both, and aim to explore the genetic history and patterns within a defined locale.
- mtDNA Lineage Projects are designed for those interested in tracing direct maternal lineages, regardless of surname changes due to marriage. These projects bring together individuals who share a common maternal heritage.
- Special Interest or Family Finder Projects focus on specific people or issues. Sometimes projects focus on a particular couple (as discovered by Family Finder autosomal testing) or a group united by historical, cultural, or genealogical interests. These can include adoptee projects and projects for descendants of notable groups such as indigenous communities, pilgrims, or nobility.
“The Group Project system allows Group Project Administrators to organize members into subgroups based on their project goals. Y-DNA and surname projects typically focus on grouping the members into genetic subgroups based on Y-SNP and Y-STR markers or based on genealogical or geographical information.” [25]
At the begining of 2023, three types of group projects represented about 92 percent of the group projects. YDNA surname projects constituted the majority of group projects. Surname projects represented roughly three quarters of the groups projects (see illustration one below). Nine percent of the group projects focus on autosomal DNA connections. About eight percent of the group projects are focused on a geographical areas for YDNA.
Illustration Two: FamilyTreeDNA Group Project Types (as of February 2023)
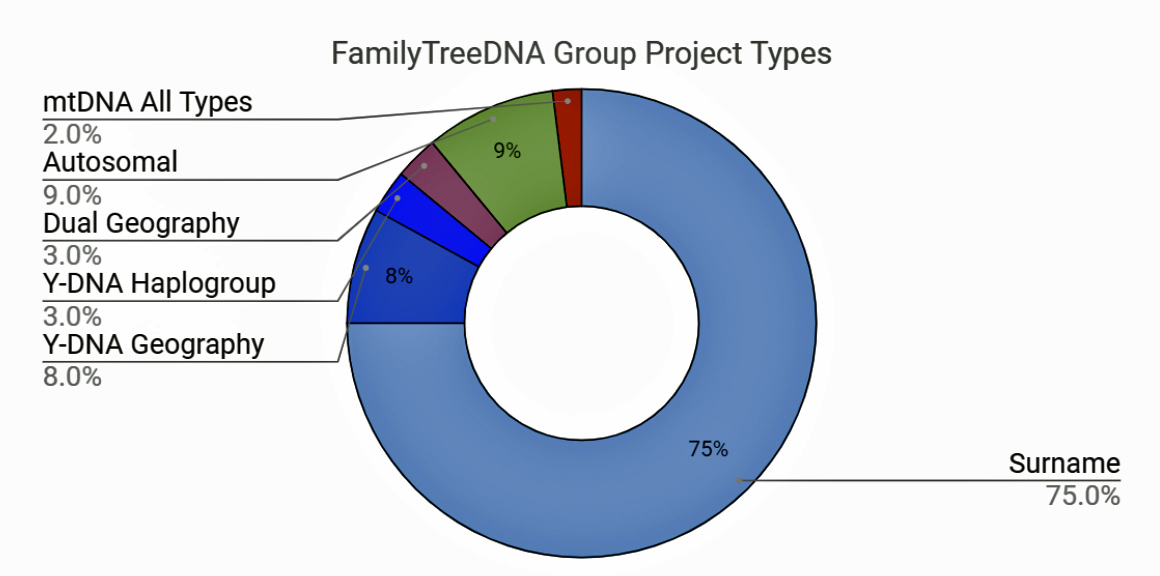
About 3 percent of the projects are what are known as ‘dual geography’ (or dual geographical) projects. They are regional projects that deliberately collect and analyze Y-DNA, mtDNA, and often autosomal (Family Finder) results together for a single country, region, or locality. [26] Another three percent of the group projects involve YDNA haplogroups.
Advantages of Joining Multiple Work Groups
Joining multiple YDNA FTDNA Work Projects maximizes your potential research return by providing a different research focus, genetic genealogical analysis and collaborative expertise that a single project cannot offer. The different research focus or perspective for each of these different work projects provide complementary benefits in genealogical research. [27]
J. David Vance’s framework of three periods of ancestry is useful for discussing the relative advantages of and coherence between each type of YDNA Work Project. The utility of YDNA evidence changes fundamentally depending on how far back in time you are looking. Three distinct temporal layers require different tools, different interpretive frameworks, and yield different types of knowledge (see illustration three). [28]
The three period framework emerged from a challenge familiar to every genealogist — the brick wall. When documentary records run dry, genealogists historically had nowhere to go. Vance’s insight was that Y-DNA does not simply confirm what records show; it can extend knowledge into temporal zones where no records exist, though the nature of that knowledge differs by period.
Genealogy – The most recent period where generations of named ancestors have been documented through traditional records research (birth, marriage, death records, censuses, etc.), possibly corroborated by DNA testing. This is the era of the documented family tree.
Period of Lineages (or Clans) – The intermediate period beyond traditional “brick walls” where specific named ancestors cannot be identified, but surname lineages or clans can be traced through YDNA matching. This netherworld connects documented genealogy to deep ancestry.
Deep Ancestry – The most ancient period reaching back thousands of years, traced through haplogroups and ancient migration patterns (e.g., Mesolithic hunter-gatherers, Neolithic farmers, Bronze Age steppe populations). This reveals prehistoric origins and continental migrations.
Illustration Three: Vance’s Three Periods of Ancestry
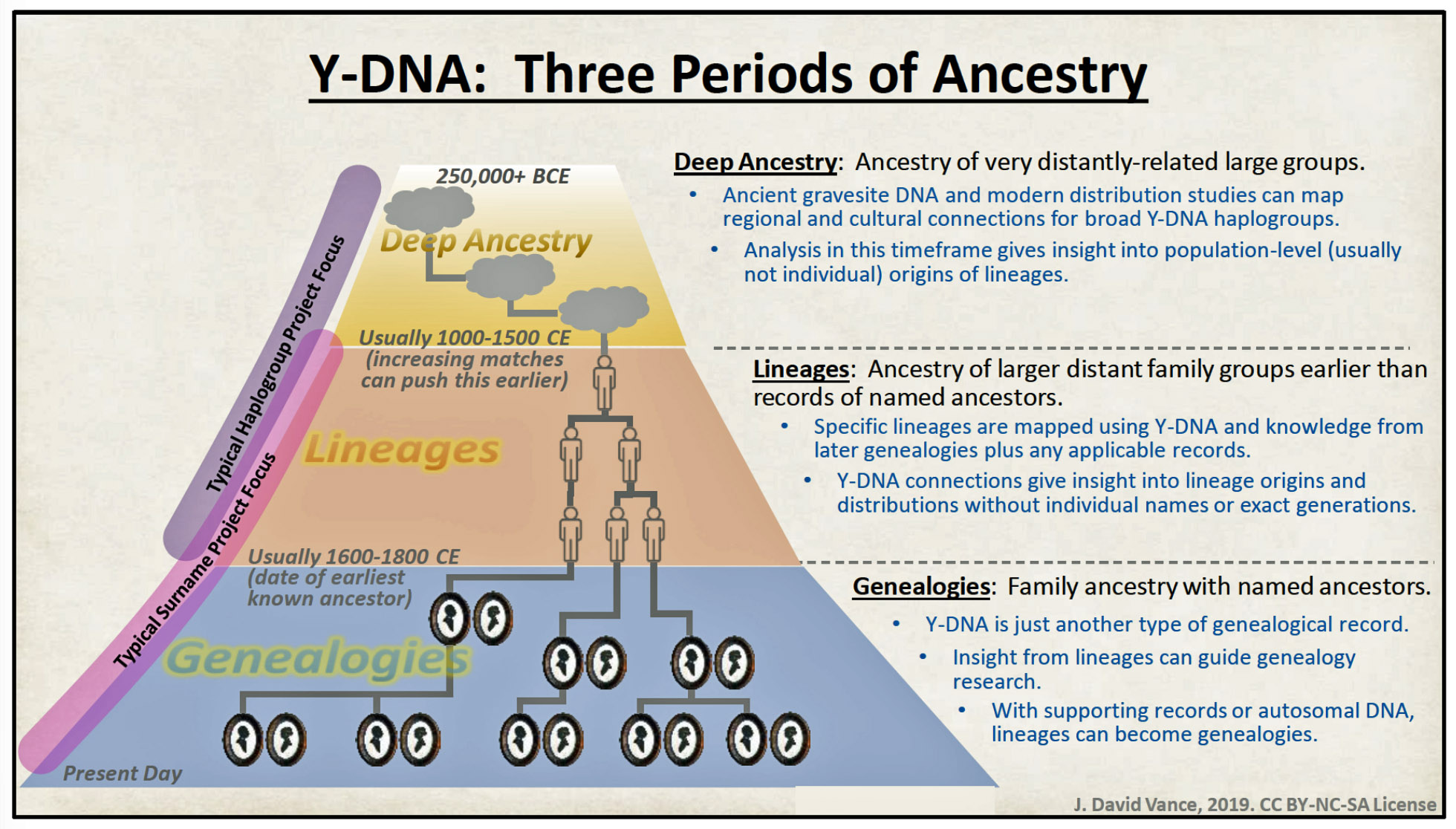
This three-tiered view allows researchers to seamlessly transition from documenting named ancestors (Genealogy) to mapping unnamed lineage ancestors (Lineages) to exploring prehistoric migrations (Deep Ancestry) within a single integrated outlook.
Vance emphasizes that the boundary between genealogy and the period of lineages is not a fixed date but shifts as new genealogical discoveries push ‘brick walls’ or the absence of information further back. DNA testing, particularly YDNA, serves as the bridge that connects the documented era to the deeper, pre-surname past.
Based on Vance’s framework and FamilyTreeDNA’s (FTDNA) work project structure, a correlation can be made between the three periods of ancestry and the utility of the types of Y-DNA Group Projects hosted on the FTDNA platform (see table four).
Table Four: Vance’s Three Period of Ancestry and FTDNA’s Project Structure
| Vance’s Period | Time Depth | Primary DNA Markers | Perdominant FTDNA Project | Project Purpose / Goals |
|---|---|---|---|---|
| Genealogy | ~1500 CE to present (documented era) | Y-STRs (37–111 markers) | Surname Projects | Connect test kits with the same surname to identify common ancestors within genealogical time; break through brick walls using STR matches. |
| Lineages | ~500–1500 CE (pre-surname to early surname era) | Y-STRs + Y-SNPs (Y-111 & Big Y-700) | Surname Projects (deep subclades) & Geographic Projects | Map unnamed ancestors through mutation history trees (MHT); identify lineage branching points and regional clusters before surnames stabilized. |
| Deep Ancestry | 500+ back to Paleolithic and beyond (prehistoric migrations) | Y-SNPs (haplogroup-defining) | Haplogroup Projects | Trace ancient migration paths, identify haplogroup origins, and correlate with archaeological cultures using the Y-DNA phylogenetic haplotree. |
My Involvement with FTDNA Projects
I originally was a member of four projects based on my general goal of expanding and tracing the family genealogy of the Griff(is)(es)(ith) family through YDNA genetic research. The four initial FTDNA project groups were:
- The GRIFFI(TH,THS,N,S,NG…etc) surname project: This project is intended, as its name indicates, to provide an avenue for exploring the genetic connections that may exist with YDNA testers that have Griffith, Griffiths, Griffin, Griffis, Griffing and other derivative surnames. [29]
- The G-L497 Working Group: This project is a large one in terms of members and in the number of project managers. It is a well developed project that includes a wealth of research links and maintenance of phylogenetic tree maps. The project includes FTDNA test kit results with the G-L497 SNP mutation. The L-497 is a major branch or subclade of the G-haplogroup that reflects the migration route into central Europe. The primary goal of the project is to identify new subgroups of haplogroup G-L497 which will provide better focus on the migration history of haplogroup G-L497 ancestors. The L497 haplogroup is part of the paternal ancestral migratory path of the Griff(is)(es)(ith) lineage. [30]
- The Welsh Patronymics project is designed to establish links between various families of Welsh origin with patronymic style surnames. [31]
- The Wales Cymru DNA project collects the DNA haplotypes of individuals who can trace their Y-DNA and/or mtDNA lines to Wales. [32]
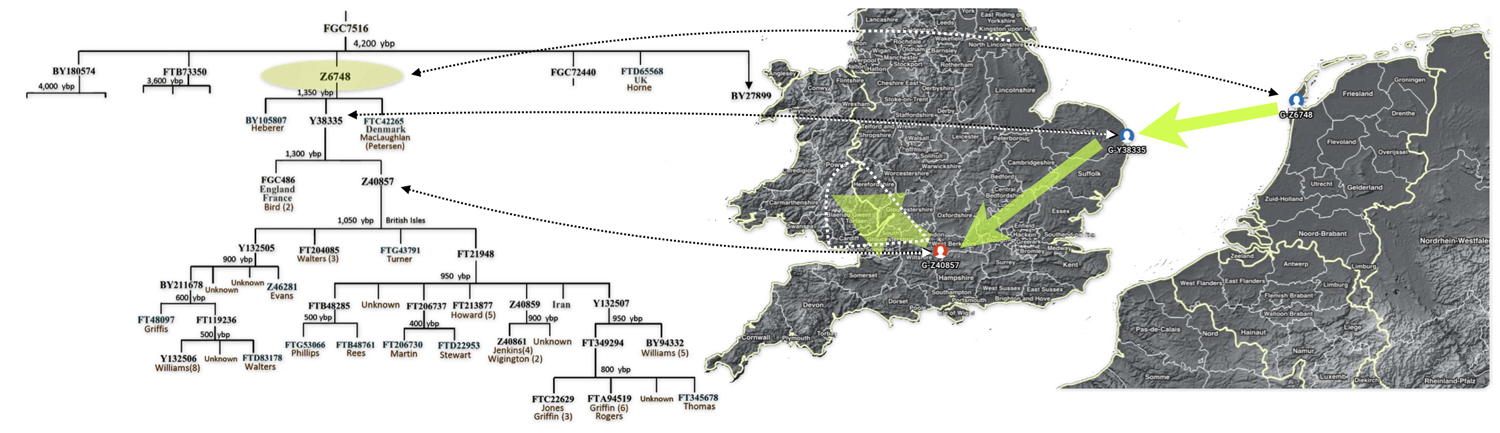
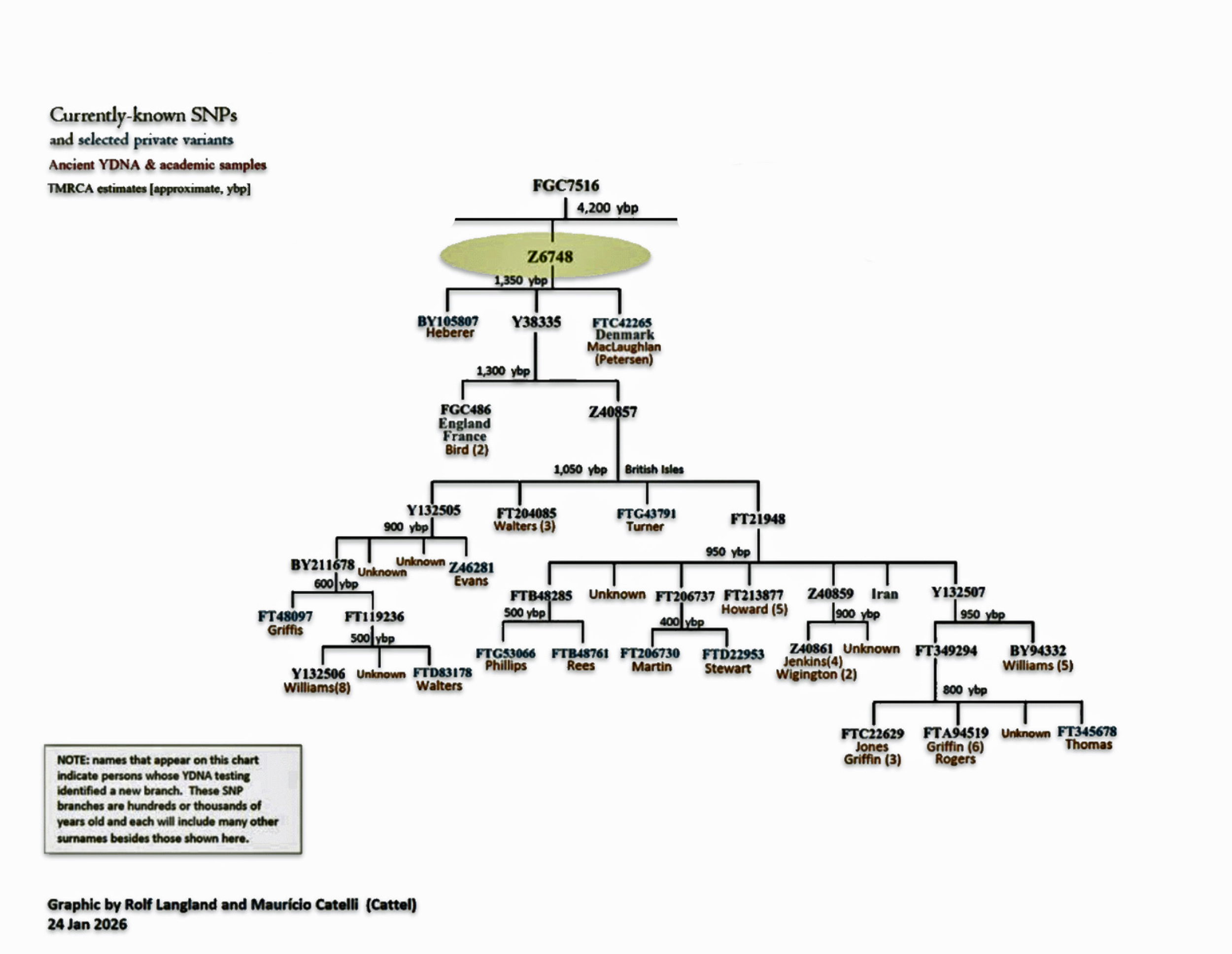
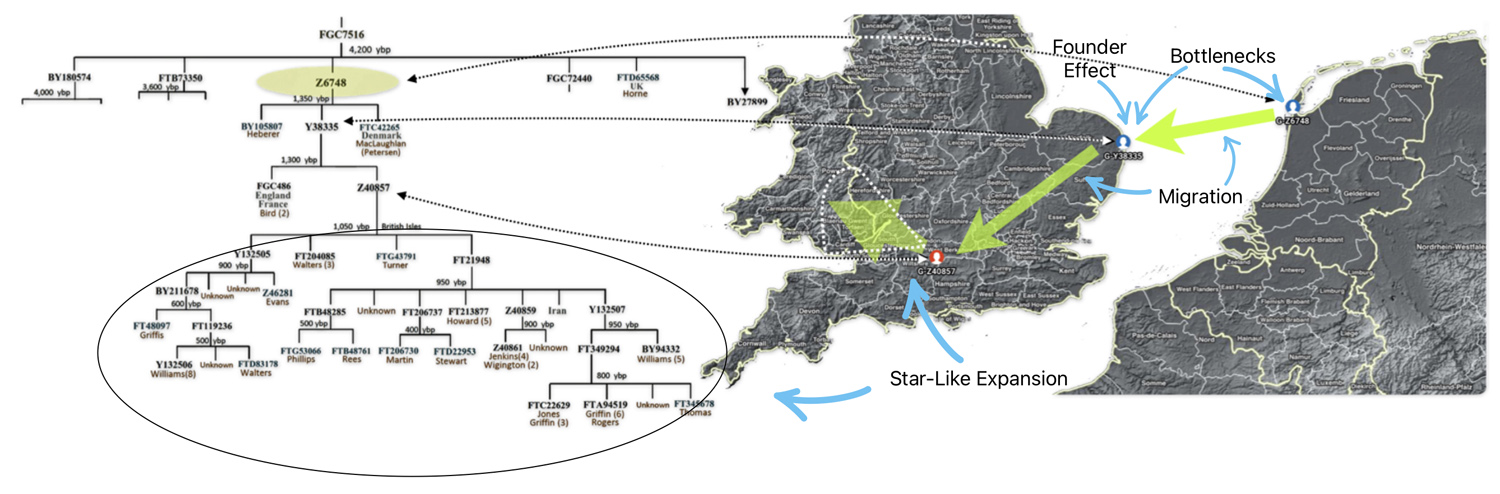
Each of these work groups provide benefits for discovering YDNA matches and specific angles for discovering patterns and relationships among group project members. My involvement with the G-L497 group project has been particularly beneficial. Since my YDNA lineage can be traced back to the L497 haplogroup, thre is a small group of YDNA testers that belong to the G-Z6748 subclade. The project administrators of the group also provide noteworthy research documents and information, such as the phylogenetic tree charts (see illustration four).
Illustration Four: Known Haplotree of G-Z6748 as of January 2026
The Inception of the G-Z6748 Haplogroup Project
In February of 2022, Thomas Weaver, one of the volunteer administrators associated with the G-L497 project created a new haplogroup project based on the descendents of G-Z6748. [31] Weaver reached out through email correspondence to targeted individuals, including me, who were members of the G-L497 project that could trace their lineage back to the G-Z6748 haplogroup.
“I am a new co-admin of the FTDNA G-L497 Haplogroup Project and have created a new FTDNA G-Z6748 Haplogroup Project to focus specifically on your UK branch.
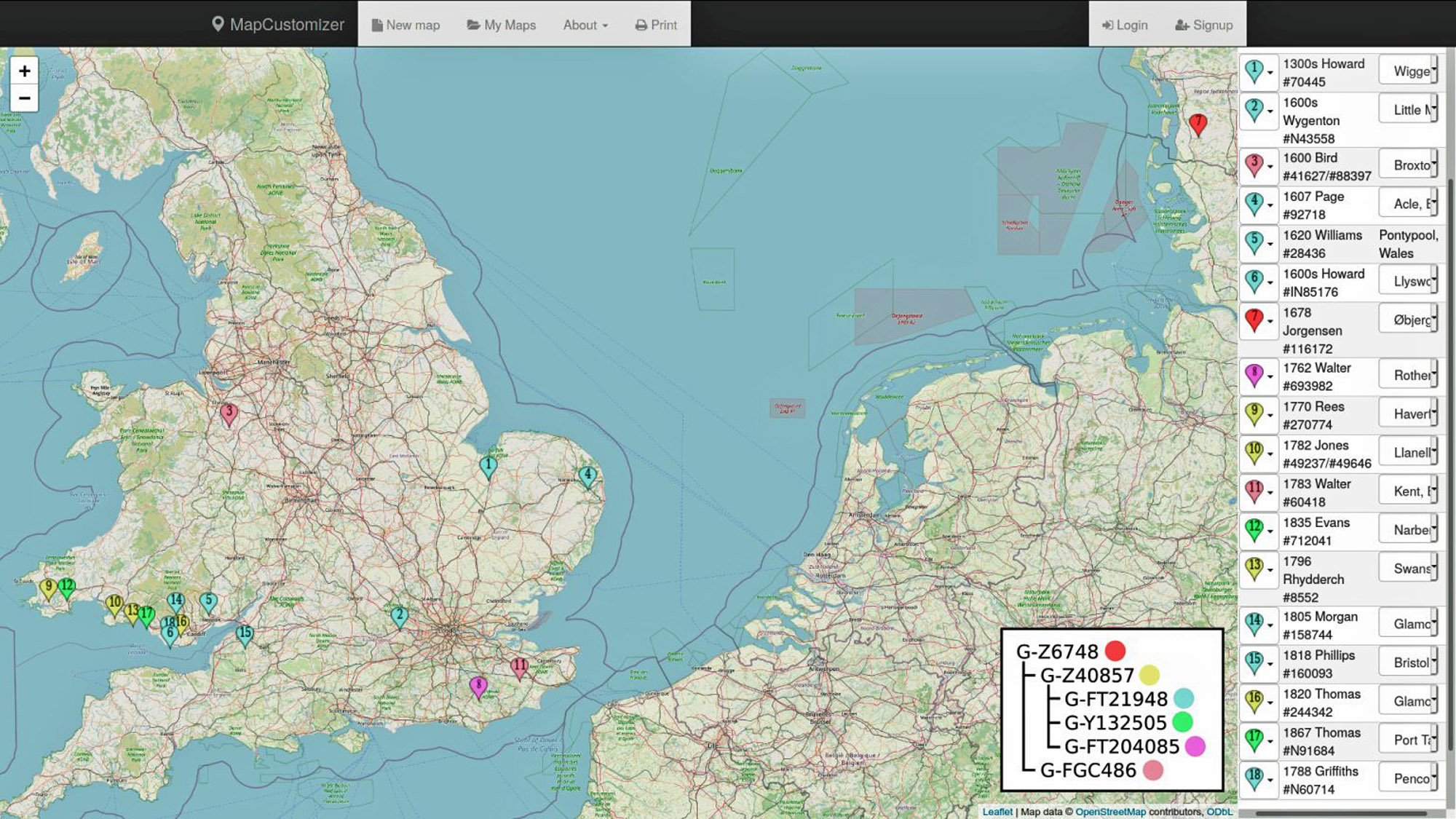
“I created the attached map (see illustration five below) that shows the towns of participants who have traced their earliest ancestor to Europe. It helps us see the homelands of the group. The common ancestor will be before surnames, probably in the Early Middle Ages, which is why we see multiple surnames.” [33]
“That map shows you the year, surname, and town of origin for each of the listed kits. . . . Most of them have different surnames than you. The common ancestor is a man from the Early Middle Ages, but most of his descendants appear to be from Southern Wales. Each of those names and markers represent independent migrant lines and where they are tracing their earliest known paternal ancestor.
“As a group, these different lines make a powerful statement as to where the group originated. Your Big Y results specifically show us your branch, and helping others see the value of upgrading will reveal the origin of each branch, refining the group’s results.” [34]
Illustration Five: Map of G-Z6748 Testers’ Self Reported Earliest Known Ancestors
The Unique Advantages of the G-Z6748 Haplogroup Work Group
The members of the G-Z6748 Work Group have a common YDNA ancestor who is related to haplogroup G-Z6748. He lived at the end of the Roman era or perhaps the late iron age / early medieval times in an area that is now known as the Netherlands. [3] Another significant fact associated with this ancestor is that at least one of his descendants migrated to what is now known as the British Isle.
The G-Z6748 Haplogroup Work Group is focused on identifying and inviting YDNA testers that have tested positive for the G-Z6748 SNP to join the research work group. In addition, an ongoing objective is to document the evolving SNP branches or subclades of the genetic descendants of the most recent common ancestors (MRCA) of this haplogroup. Another objective is to facilitate the discovery of YDNA matches among work group members.
Based on the definitions provided above for the various FTDNA work projects, the G-Z66748 work group is a downstream haplogroup work group that is largely delimited by geography. The descendants of the G-Z6748 Haplogroup can be traced on the British Island and possibly the contours of the coastal northwestern European continent.
The surnames of the modern day descendants of G-Z6748 vary. This is due to the fact that surnames emerged and became prominent in various parts of the British Isle and northwestern coastal Europe about one thousand years after this ancestor and his descendants lived.
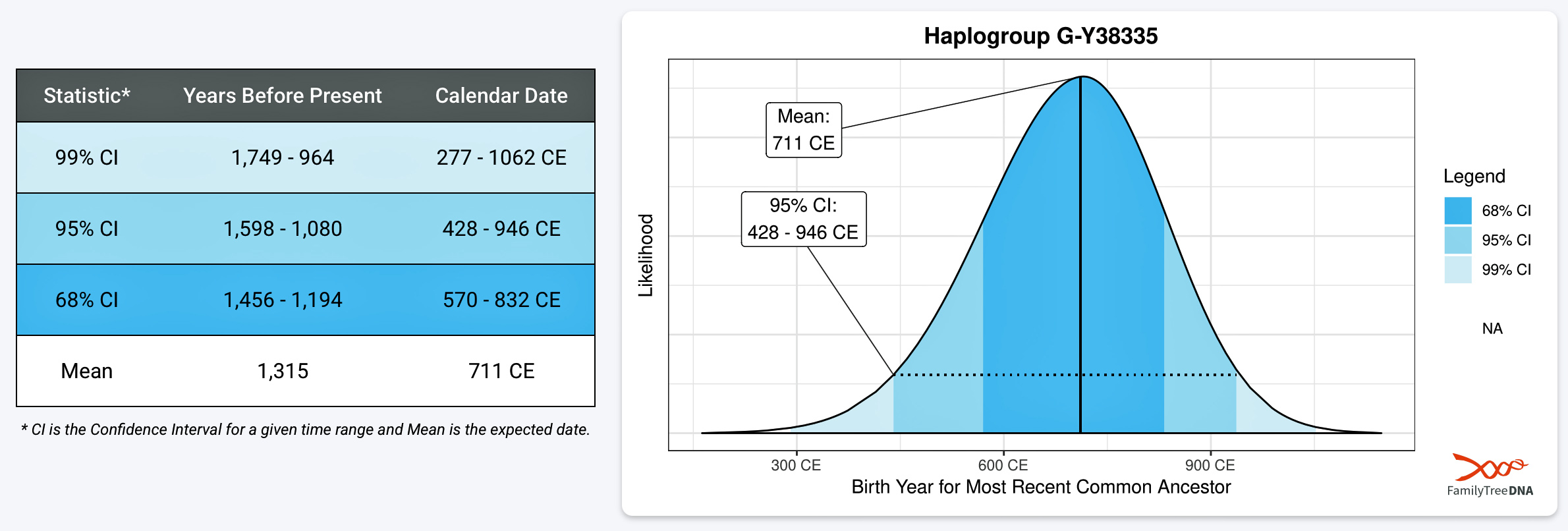
Illustration Six depicts a phylogenetic tree of descendants of the G-Y38335 haplogroup. This is a descendant of the G-Z6748 haplogroup – the first ancestors to migrate to the British Isle. The tree was generated through the use of a computer program created by David Vance. [35] Added to the tree diagram is a shaded area that depicts approximately when surnames emerged in relation to the haplogroup subclade. [36] This is a practical illustration of Vance’s boundary between genealogy and the period of lineages
Illustration Six: The Descendants of G-Y38335 and the Emergence of the Use of Surnames
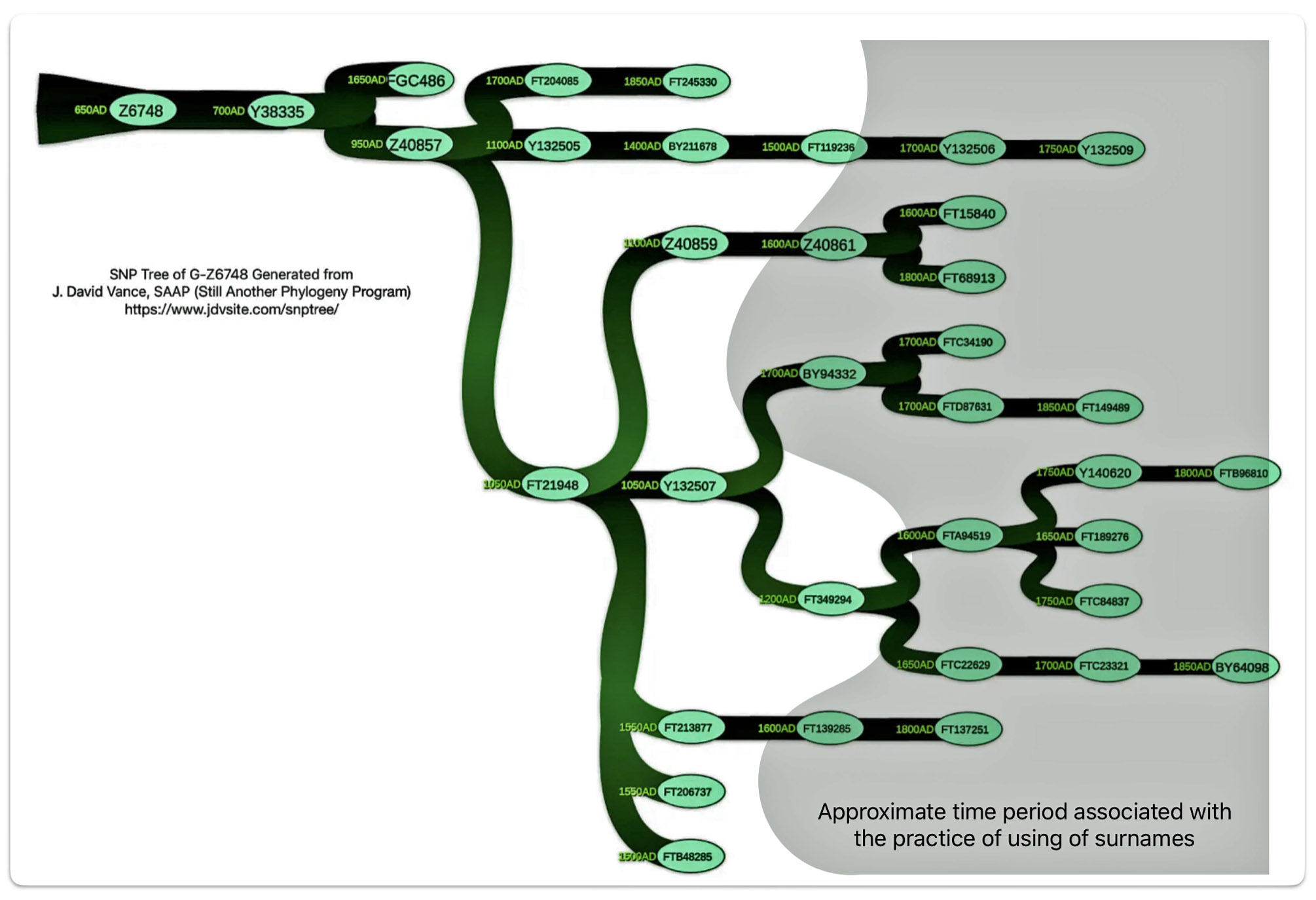
The resultant historical effect is that many of the documented genetic descendants have different surnames – reflecting names such as Williams, Griffis, Griffith, Griffin, Jones, Jenkins, Howard, and Wigington, among others.
A unique advantage of being a member of this group as well as being a member of other surname groups is it prevents the error of assuming a surname equals a single genetic lineage (possible in surname projects) while also avoiding the noise of analyzing regional data without a specific lineage anchor (a risk in geographical projects).
A downstream haplogroup project, such as G-Z6748, functions as a “regional filter” that allows you to see your Y-DNA lineage in the context of deep time and landscape, rather than just the last 500 years of surname usage.
Table Five: Advantages of Being a Member of G-Z6748 Haplogroup
| Mechanism | How It Identifies Migration and ‘Clans’ |
|---|---|
| Pre-Surname Context | Surnames are recent (medieval/modern), but clans often moved millennia earlier. Geographical projects group by region, allow one to see haplogroup branches movement before surnames existed. |
| Cross-Surname Correlation | Unlike surname projects, geographical and downstream haplogroup projects include all paternal lines from a region. This reveals if multiple distinct surnames (e.g., Griffith, Griffis, Williams, etc) share a recent common ancestor, indicating a founder effect or a single clan that fractured and adopted different surnames. |
| Cluster Analysis | Administrators group members by STR signatures and terminal SNPs specific to the region and subclades of G-Z6748. If a clan’s specific subclade forms a tight cluster but is scattered in a broader geographic project, it signals a localized settlement event. |
| Ancient DNA Integration | Tools like Globetrekker (powered by FTDNA’s Discover™) integrate ancient DNA samples and Least Cost Path (LCP) modeling to visualize exactly how your clan’s ancestors likely moved across the landscape. |
| Outlier Detection | By comparing specific SNP branches against a larger geographic regional baseline and othe subclades, administrators can spot “outliers” that indicate specific migration events (e.g., the impact of the Normal invasion). |
Overlapping Membership in FTDNA YDNA Projects is Good for Everyone Involved
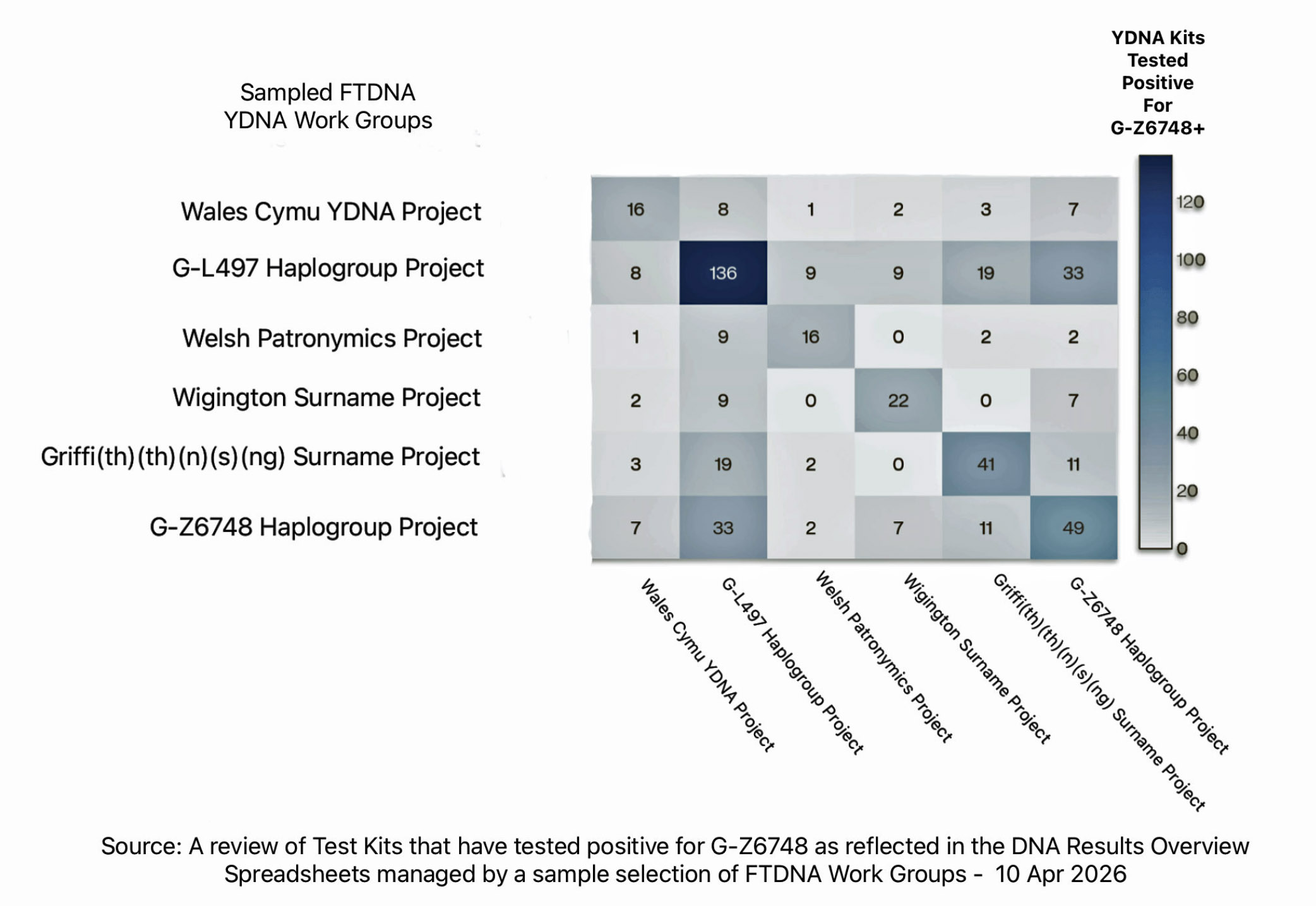
To date, there are approximately 150 YDNA testers that are documented to be modern descendants of the most recent common ancestor of haplogroup G-Z6748. That number may go up as more male individuals are tested. [37]
Joining FTDNA work groups is voluntary. The G-Z6748 Work Group has about 50 of those 150 test kits in the work group. The upstream G-L497 ‘parent’ Haplogroup work group has the largest number of test kits that have tested positive for the G-Z6748 SNPs associated with the G-Z6748 haplogroup. Other surname and geographical work groups have G-Z6748+ test kits ranging from about 16 to 40 YDNA testers. Some of these test kits overlap in each of the sampled work groups (see illustration seven).
Illustration Seven: FTDNA YDNA Testors that have Tested Positie for G-Z6748 and their distribution in FTDNA Work Groups
Joining multiple FTDNA work groups not only can aid individuals associated with specific test kits but also benefits each of the individual work groups.
Table Six: Key Advantages of Joining Multiple FTDNA Projects
| Advantage | How It Helps Your Research |
|---|---|
| Multi-Scale Context | Comparing your results in a Surname Project (recent genealogy), a Haplogroup Project (deep ancestry), and a Geographical or Downstream Haplogroup Project (regional migration and lineages) has potential to reveal patterns and connections that would not be obvious in isolation. |
| Guidance | Different project administrators specialize in different areas (e.g., a specific surname vs. a broad haplogroup, providing targeted advice. |
| Enhanced Matching | Projects utilize the Y-DNA Results Overview Report, allowing administrators to group you with close matches based on STR signatures that might be missed in general or other group project’s matching . |
| Upstream and Downstream Collaboration | Joining “upstream” or “downstream” haplogroup projects allows your specific surname data to contribute to broader population studies, often leading to the discovery of new SNPs that refine your branch. |
Sources
Feature Image: The banner image consistes of two images. The image on the left is a modifed version of a Phylogenetic Tree created by Rolf Langland and Maurício Catelli (see reference below). The image on the left is a map I have created that illustrates the estimated migratory path between the MRCA of G-Z6748 and G-Y38335 and G-Z40857.
Feature Image source for map: The source for creating the map is based on a variety of historical and archaeological studies as well as the estimates derived from the FTDNA Globetrekker tool, see Jim Griffis, Migrating to East Anglia, March 31, 2026, Griffis Family: Selected Stories from the Past, https://griffis.org/migrating-to-east-anglia/
Feature Image source for phylogenetic tree: Rolf Langland and Maurício Catelli, G-FGC477 / Chart D – v6 – 2 pages (Jan 26), G-L497 Y-DNA Work Project, https://drive.google.com/file/d/1U_-FfascgkP2kS4nVPEPQVr0l6w8Qc7U/view
[1] Quote: Haplogroup G-M201, Wikipedia, This page was last edited on 20 February 2026, https://en.wikipedia.org/wiki/Haplogroup_G-M201
The quote references the ISOGG haplogroup G2a3b1 which is G-P303 . A direct match to G2a3b1 is not found, but 3 steps up the haplotree is G2a which has an equivalent name of G-P15.
See also:
Haplogroup G-P303, Wikipedia, This page was last edited on 26 January 2026, https://en.wikipedia.org/wiki/Haplogroup_G-P303
“There are seeming pockets of unusual concentrations within Europe. In Wales, a distinctive G2a3b1 (G-P15) type (DYS388=13 and DYS594=11) dominates there and pushes the G percentage of the population higher than in England.“
DYS399 and DYS594 stand for DNA Y-chromosome Segments. They are specific short-tandem repeat (STR) markers located on the Y-chromosome used in genetic genealogy to trace paternal ancestry. DYS markers, designated by the HUGO Gene Nomenclature Committee, identify specific spots where DNA sequences repeat, helping men determine relatedness to others through their direct paternal line.
Key Details About DYS Markers (e.g., DYS399 and DYS594):
- Paternal Tracking: DYS markers only exist on the Y-chromosome, passing from father to son with few changes, making them ideal for surname projects and genealogical research.
- STR (Short Tandem Repeat): These markers measure the number of times a short DNA sequence repeats, such as GATA-GATA-GATA (3 repeats).
- Mutation Rates: While highly stable, these markers can mutate, allowing researchers to estimate the time to the most recent common ancestor (TMRCA) between two men.
- Component of Y-DNA Profiles: Results for DYS399, alongside others like DYS390 or DYS393, form a Y-STR haplotype profile.
Understanding the Admin – Y-DNA Results Overview Report, FamilyTreeDNA, https://help.familytreedna.com/hc/en-us/articles/11165708791311-Understanding-the-Admin-Y-DNA-Results-Overview-Report#h_01JBYS1DRY1CMCC0FVK83ER1GQ
A review of the DYS values for 399 and 594 for members of the G-Z6748 FamilyTree Project confirms this observation. The following is the G-Z6748 – Y-DNA Results Overview for the FamilyTreeDNA project. As reflected in the chart, the value for DYS399 is 13 for all members. The value for all but one member for DYS594 is 11.
G-Z6748 – Y-DNA Results Overview (as of April 2026)

[2] Haplogroup P-303, Wikipedia, This page was last edited on 26 January 2026, https://en.wikipedia.org/wiki/Haplogroup_G-P303
[3] The map is from an innovative study that systematically assessed the association between genetic variation in the male-specific region of the Y chromosome (MSY) and cardiovascular disease outcomes. The researchers conducted a kin-cohort analysis of family disease history using the largest sample to date. The study involved testing 90 MSY haplogroups against several cardiovascular health indicators including coronary artery disease, hypertension, blood pressure, classical lipid levels, and all-cause mortality.
The primary finding of the study was that their models showed little evidence for an effect of any MSY haplogroup on cardiovascular risk in participants. An important secondary finding was that Y chromosome haplogroups carried by White British individuals demonstrate strong geographic structuring across Great Britain. The researchers observed that certain lineages are more prevalent in specific regions.
The Timmers and Wilson haplogroup maps offer genealogical researchers several distinctive advantages beyond typical commercial DNA project maps. With 152,186 unrelated white British men, this is the largest Y chromosome geographic survey ever conducted for Britain—far exceeding commercial projects like FamilyTreeDNA’s British Isles Project (typically thousands of participants). This scale provides:
- Statistical robustness for rare haplogroups that appear sporadically in smaller datasets;
- Fine-grained resolution at ward and electoral division levels (the smallest UK census units), not just counties or regions; and
- Reliable frequency estimates even for subclades with only hundreds of carriers.
Unlike commercial maps showing “earliest known ancestor” pins (which suffer from recall bias and uneven sampling), these maps use official 2011 UK Census boundaries with strict inclusion criteria (minimum 100 individuals per area). This means:
- Researchers can directly correlate haplogroup distributions with historical census data, parish records, and surname distributions;
- Frequencies are population-based, not volunteer-based; and
- Geographic units are hierarchical and comparable (wards → local authorities → regions → nations).
Timmers, Paul RHJ; Wilson, James F. (2022). Prevalence of Y chromosome haplogroups by area of birth in UK Biobank, [image]. University of Edinburgh. https://doi.org/10.7488/ds/3472.https://datashare.ed.ac.uk/handle/10283/4450
[4] Griffis, Jim, Y-DNA and the Griffis Paternal Line Part Three: The One-Two Punch of Using SNPs and STRs,, February 23, 2023, https://griffis.org/y-dna-and-the-griffis-paternal-line-part-three-the-one-two-punch-of-using-snps-and-strs/
[5] Bates, Sarah, Base Pair, April 27, 2026, National Human Genome Institute, https://www.genome.gov/genetics-glossary/Base-Pair
Base Pair, Wikipedia, This page was last edited on 9 April 2026, https://en.wikipedia.org/wiki/Base_pair
[6] Estes, Roberta, STRs vs SNPs, Multiple DNA Personalities, 10 Feb 2014, DNAeXplained – Genetic Genealology, https://dna-explained.com/2014/02/10/strs-vs-snps-multiple-dna-personalities/
Estes, Roberta, STRs and SNPs – Are STR Markers Still Useful for Y DNA?, 3 Dec 2021 , DNAeXplained – Genetic Genealology, https://dna-explained.com/2021/12/03/strs-and-snps-are-str-markers-still-useful-for-y-dna/
Li M, Zhang H, Tao R, Chen A, Zhou P, Yu C, Bian Y, Zhang S, Fang C, Li C. Exploring Y-chromosomal STRs and SNPs for forensic and genetic insights in the Jiangsu Han population. BMC Genomics. 2025 May 2;26(1):440. doi: 10.1186/s12864-025-11634-6. PMID: 40316924; PMCID: PMC12048932. https://pmc.ncbi.nlm.nih.gov/articles/PMC12048932/
Y-DNA tools, International Society of Genetic Genealology Wiki, This page was last edited on 8 February 2026, https://isogg.org/wiki/Y-DNA_tools
[7] Microsatellite, 27 Apr 2026, National Human Genome Institute, https://www.genome.gov/genetics-glossary/Microsatellite
Y-STR Results Frequently Asked Questions, FamilyTreeDNA Help Center, Page accessed 12 Apr 2026, https://help.familytreedna.com/hc/en-us/articles/4408071453711-Y-STR-Results-Frequently-Asked-Questions
[8] Single Nucleotide Polymorphisms (SNPS), (SNPS) , 27 Apr 2026, National Human Genome Institute, https://www.genome.gov/genetics-glossary/Single-Nucleotide-Polymorphisms-SNPs
Estes, Roberta, STRs vs SNPs, Multiple DNA Personalities, 10 Feb 2014, DNAeXplained – Genetic Genealology, https://dna-explained.com/2014/02/10/strs-vs-snps-multiple-dna-personalities/
Estes, Roberta, STRs and SNPs – Are STR Markers Still Useful for Y DNA?, 3 Dec 2021 , DNAeXplained – Genetic Genealology, https://dna-explained.com/2021/12/03/strs-and-snps-are-str-markers-still-useful-for-y-dna/
[9] Estes, Roberta, STRs vs SNPs, Multiple DNA Personalities, 10 Feb 2014, DNAeXplained – Genetic Genealology, https://dna-explained.com/2014/02/10/strs-vs-snps-multiple-dna-personalities/
[10] Y chromosome DNA haplogroup, International Society of genetic Genealology, https://isogg.org/wiki/Y_chromosome_DNA_haplogroup
Human Y-chromosome DNA haplogroup, Wikipedia, This page was last edited on 12 April 2026, https://en.wikipedia.org/wiki/Human_Y-chromosome_DNA_haplogroup
Estes, Roberta, Y DNA: Part 2 – The Dictionary of DNA, 27 Jan 2020, DNAeXplained – Genetic Genealology, https://dna-explained.com/2020/01/27/y-dna-part-2-the-dictionary-of-dna/
[11] In database, software, and genetic contexts, Y-DNA haplogroups are structured as recursive sets and nested hierarchies (also known as a directed acyclic graph or phylogenetic tree. These trees map the evolutionary history of paternal lineages, where each haplogroup is defined by a specific single nucleotide polymorphism (SNP) mutation that occurred at a specific time and place
Key Concepts in Haplogroup Hierarchies:
- Nested Structure: A haplogroup (e.g., R1b1a1a2) is a subset of a broader, more ancestral group (e.g., R1b1a1a), which in turn is a subset of an even broader group (e.g., R). As you move “down” the tree (towards more recent times), you are navigating into more specific subclades.
- Phylogenetic Trees (Y-DNA): These trees demonstrate relationships between Y-chromosome lineages, starting with broad “backbone” haplogroups and branching into over 90,000 sub-branches (subclades) in specialized databases like FamilyTreeDNA.
- Nomenclature: Haplogroups are often labeled using a nested nomenclature system with numbers and letters identifying sublineages.
- Database Management: Software systems (like ISOGG or Yleaf) use tree traversal algorithms to manage and assign haplogroup labels, navigating this hierarchical structure to classify new Y-DNA sequences.
Y Chromosome Consortium. A nomenclature system for the tree of human Y-chromosomal binary haplogroups. Genome Res. 2002 Feb;12(2):339-48. doi: 10.1101/gr.217602. PMID: 11827954; PMCID: PMC155271. https://pmc.ncbi.nlm.nih.gov/articles/PMC155271/
Haplogroup, Wkipedia, This page was last edited on 7 January 2026, https://en.wikipedia.org/wiki/Haplogroup
Rowe-Schurwanz, Katy, 2 Jul 2024, Interpreting Y-DNA Test Results: Y-DNA Haplogroups, FamilyTreeDNA Blog, https://blog.familytreedna.com/interpreting-y-dna-test-results-haplogroups/
FamilyTreeDNA’s Y-DNA Haplotree: 90,000 Branches and Counting, FamilyTreeDNA Blog, https://blog.familytreedna.com/ydna-haplotree-90000-branches
[12] My detailed haplogroup path is the following: G-M201> L89> L156> P15> L1259> L30> L141> P303> L140> PF3346> Z3065> PF3345> L497> CTS9737> Z1900> Z6901> Z1817> Z727> FGC477> FGC7516> Z6748> Y38335> Z40857> Y132505> BY211678
Ancestral Path of G-BY211678, FamilyTreeDNA, https://discover.familytreedna.com/y-dna/G-Z6748/path
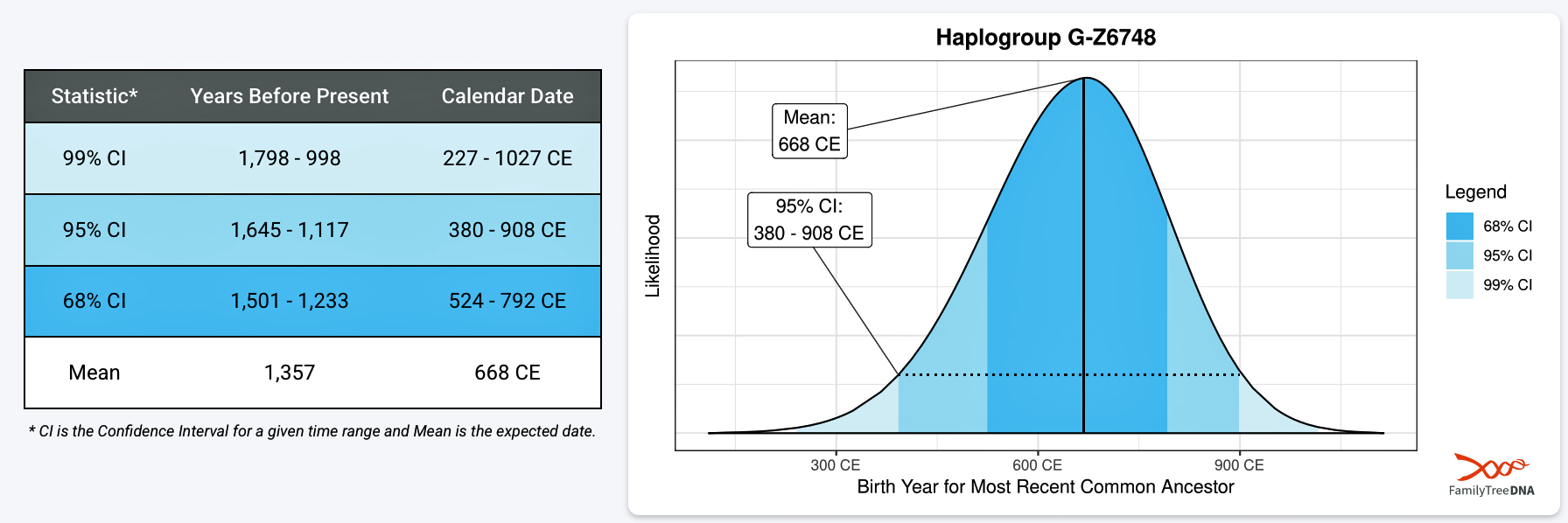
[13] The man who is the most recent common ancestor of this line is estimated to have been born around 650 CE. He is the ancestor of at least 2 descendant lineages known as G-Y38335 and 1 yet unnamed lineage.
Your Haplogroup Story: G-Z6748, FamilyTreeDNA, https://discover.familytreedna.com/y-dna/G-Z6748/story
[14] The 29 SNPs that are associated with the G-Z6748 branch are: G-Z6748, BY8142, FGC476, FGC479, FGC481FGC482,FGC483, FGC484, FGC485, FGC487, FGC488, FGC490, FGC496, FGC498, FGC499, FGC500, FGC502, FGC504, FGC505, FGC506, FGC507, FGC509,FGC511, FGC512, FGC516FGC517,,FGC518, FT73641, and Y172988
Scientific Details of G-Z6748: Variants, FamilyTreeDNA, Accessed 3 Jun 2026, https://discover.familytreedna.com/y-dna/G-Z6748/scientific?section=variants
[15] Ancestral Path of G-BY211678, FamilyTreeDNA, https://discover.familytreedna.com/y-dna/G-Z6748/path

Estes, roberta, Glossary – Terminal SNP, 29 Nov 2017, DNAeXplained, https://dna-explained.com/2017/11/29/glossary-terminal-snp/
[16] A novel SNP is a previously unreported genetic variant. It means this specific mutation at a specific location on the genome has not been documented in public genetic databases. A novel variant is simply “new to science.” It does not mean it is unique it merely has not been mapped yet.
As of the date of this story, based on an YFull analysis, I have seven novel or private variant SNPS. One of which, FT48097, is verified by Sanger sequencing YSseq 40 by Yull.com
Novel SNPs Associated with my Y700 Test Results Based on Analysis by YFull
| Name | Position Hg38 | Reference | Derived | Q | Qual | Reads | T2T Only |
|---|---|---|---|---|---|---|---|
| Y172952 | 7246246 | C | T | 100 | Best qual | 8 | – |
| FT48097 | 15863570 | C | T | 100 | Best qual | 29 | – |
| 5357260 | T | C | 100 | Ambiguous qual | 2 | – | |
| 13953027 | A | T | 1 read | One reading! | 1 | – | |
| 15064752 | T | C | 1 read | One reading! | 1 | – | |
| 15531370 | T | C | 1 read | One reading! | 1 | – | |
| 20693374 | G | A | 1 read | One reading! | 1 | – |
Terminal SNPs for BY211678YFull Y-Chr Sequence Interpretation Service, YFull, https://www.yfull.com/snp/private/
Here is what each YFull private SNP table heading means in that context:
- Position Hg38: Genomic coordinate of the variant on the Y chromosome in the GRCh38/hg38 human reference assembly, i.e. the base position on the current standard reference genome rather than hg19/GRCh37.
- Reference (TACG): The reference-base state at that position in hg38, using standard nucleotide codes T, A, C, or G. In other words, this is the allele present in the reference genome sequence before any sample-specific mutation.
- Derived: The non‑reference allele that YFull has called for your sample at that position, i.e. the SNP variant relative to the reference sequence. This is the putative “mutated” state on your Y line; it is what makes the site a private/novel SNP for you or your clade.
- Q: Per‑variant quality score that summarizes confidence in the variant call, usually on a Phred‑like scale derived from the underlying read and mapping qualities. Higher values mean stronger statistical support that the derived allele is real and not a sequencing or alignment error.
- Qual: Per‑read or per‑site base quality metric as reported from the mapper/caller, again Phred‑scaled; this reflects how reliable the individual base calls are at that position in your BAM/CRAM, and is one of the ingredients used by YFull when assigning their own Q and star rating. Very low Qual values flag positions where the chemistry or base‑calling is noisy, even if there are multiple reads.
- Reads: Number of sequencing reads from your sample that cover this genomic position, typically counting reads that support either the reference or the derived allele (YFull may separately track how many support the derived state when you click into the detail view). SNPs with only one or two reads are generally considered unreliable and may be excluded from age estimation or tree placement.
- T2TOnly: Indicator that this position or allele falls within sequence that exists in the complete Telomere‑to‑Telomere (T2T‑CHM13) assembly but is absent, incomplete, or problematic in hg38, or that the confident call relies on mapping against T2T rather than hg38 alone.
Swords, Christina, How to use the YFull Platform – A Tutorial for Beginners, Nebula Genomics, https://nebula.org/blog/yfull-tutorial/
Genome assembly GRCh38, The Human Genome Project, currently maintained by the Genome Reference Consortium (GRC), National Library of Medicine, https://www.ncbi.nlm.nih.gov/datasets/genome/GCF_000001405.26/
Frequently Asked Questions, Genome Reference Consortium, https://www.ncbi.nlm.nih.gov/grc/help/faq/
Reference Genome, Wikipedia, This page was last edited on 2 February 2026, https://en.wikipedia.org/wiki/Reference_genome
Serverine Catreux, Fred Farrell, Rami Mehio, Lisa Murray, Gavin Parnaby, Cooper Roddey, Mike Ruehle , Demystifying the versions of GRCh38/hg38 reference genomes, how they are used in DRAGEN and their impact on accuracy, published December 9, 2021, Illumina, https://www.illumina.com/science/genomics-research/articles/dragen-demystifying-reference-genomes.html
[17] FamilyTreeDNA, Wikipedia, This page was last edited on 14 November 2025 , https://en.wikipedia.org/wiki/FamilyTreeDNA
[18] The following is a comparative table of DNA database sizes by test type and company, based on the most recent data when the story was written (2025–2026):
DNA Database Size Comparison by Test Type
Key Takeaways:
Sources for the tables:
Peruncic, Kristina, 8 Best DNA Test Kits in 2026 (Ancestry, Health, and More),4 Mar 2026, DNAWeekly, https://www.dnaweekly.com
Peruncic, Kristina , FamilyTreeDNA vs. Ancestry 2026: Which DNA Test is Best?, 4 Mar 2026, DNAWeekly, https://www.dnaweekly.com/blog/familytreedna-vs-ancestry/
Southard, Diahan, Best DNA Test for Genealogy, 1 Dec 2025, Your DNA Guide, https://www.yourdnaguide.com/ydgblog/best-dna-tests-ancestry
MCDowell, Martin, How big is the FamilyTreeDNA database?, 14-15 Feb 2020, Genetic Genealology Ireland, https://ggi2013.blogspot.com/2020/02/how-big-is-familytreedna-database.html
Larkin, Leah, Database Sizes—March 2026, 14 Mar 2026, The DNA Geek, https://thednageek.substack.com/p/database-sizesmarch-2026
Hill, Richard, The Best DNA Testing Companies, DNA Favorites, 22 Apr 2026, https://www.dnafavorites.com/best-dna-testing-companies.html
FamilyTreeDNA, Wikipedia, This page was last edited on 16 April 2026, https://en.wikipedia.org/wiki/FamilyTreeDNA
Ancestry.com, Wikipedia, This page was last edited on 27 March 2026, https://en.wikipedia.org/wiki/Ancestry.com
Myheritage, Wikipedia, This page was last edited on 3 April 2026, https://en.wikipedia.org/wiki/MyHeritage
23andMe, Wikipedia, This page was last edited on 13 April 2026, https://en.wikipedia.org/wiki/23andMe
LivingDNA, Wikipedia, This page was last edited on 4 March 2026, https://en.wikipedia.org/wiki/Living_DNA
GEDMatch, Wikipedia, This page was last edited on 14 November 2025, https://en.wikipedia.org/wiki/GEDmatch
FamilyTreeDNA vs. 23andMe: A detailed comparison of genetic testing services, 1 Jan 2025, Nucleus, https://mynucleus.com/blog/family-tree-dna-vs-23andme
Russell, Judy, Building that mtDNA database, 30 May 2021, , The Legal Genealogist, building-that-mtdna-database
The World’s Largest mtDNA Haplotree, 14 Apr, 2026, FamilyTreeDNA Blog, https://blog.familytreedna.com/largest-mtdna-haplotree/
FamilyTreeDNA’s Y-DNA Haplotree: 90,000 Branches and Counting, 1 May 2025, FamilyTreeDNA Blog, https://blog.familytreedna.com/ydna-haplotree-90000-branches/
The World’s Largest Y-DNA Haplotree, 1 Apr 2026, FamilyTreeDNA Blog, https://blog.familytreedna.com/largest-y-dna-haplotree/
[19] CLIA-certified and CAP-accredited laboratories are clinical laboratories that meet both the federal standards from the Clinical Laboratory Improvement Amendments (CLIA) and the more stringent, voluntary requirements set by the College of American Pathologists (CAP). This dual certification indicates a high level of quality, accuracy, and reliability in laboratory testing, as CAP accreditation is considered to meet and often exceed the federal CLIA requirements.
CLIA (Clinical Laboratory Improvement Amendments)
- What it is: Federal regulations that establish quality standards for all U.S. laboratories that test human samples for health assessment.
- Purpose: To ensure the accuracy and reliability of diagnostic testing and to safeguard patient privacy.
- Oversight: Administered by a partnership of the FDA, CDC, and CMS.
- Requirement: Laboratories must be CLIA-certified to accept human samples for testing.
Clinical Laboratory Improvement Amendments (CLIA), 17 Jul 2023 U.S. Food and Drug Administraiton, https://www.fda.gov/medical-devices/ivd-regulatory-assistance/clinical-laboratory-improvement-amendments-clia
CAP (College of American Pathologists) Accreditation
- What it is: A voluntary accreditation program that is internationally recognized for its high standards.
- Purpose: To ensure that laboratory test results are accurate and reliable by assessing adherence to rigorous scientific and quality standards.
- Oversight: Conducted by the CAP, which uses its own extensive checklists and detailed requirements that are updated annually.
- Requirement: Laboratories must meet the standards of their own accreditation, which include detailed requirements for things like test validation, quality control, and proficiency testing.
- Relationship to CLIA: CAP accreditation fulfills all federal CLIA certification requirements, and laboratories that are CAP-accredited automatically meet the CLIA standards.
College of American Pathologists, Wikipedia, This page was last edited on 11 November 2025, https://en.wikipedia.org/wiki/College_of_American_Pathologists
“At FamilyTreeDNA, we value and prioritize your privacy and the security of your data as much as you do. Rest assured that we have extensively invested in safeguarding your account and personal information through multiple layers of encryption. Additionally, we take pride in owning and operating our own lab, which ensures that all testing is conducted in our CLIA-certified, CAP-accredited laboratory based in the United States.“
Ensuring your privacy & protection in our in-house lab, FamilyTreeDNA, https://www.familytreedna.com/
[20] Autosomal DNA tests are the most popular genealogy tests that analyze 22 pairs of chromosomes (autosomes) inherited from both parents, providing a comprehensive, gender-neutral overview of recent ancestry. Key uses include finding relatives (first through tenth cousins), determining ethnic percentages, and identifying genetic health markers.
Rowe-Schurwanz, Katy, What is Autosomal DNA? Beginner Guide to DNA Inheritance, 9 Apr 2026, FamilyTreeDNA Blog, https://blog.familytreedna.com/what-is-autosomal-dna/
[21] Y-DNA testing analyzes two main marker types on the Y-chromosome: STRs for recent paternal ancestry and SNPs for ancient lineage. STRs (Short Tandem Repeats) are fast-mutating markers (37–700+ tested) are used to find close matches and surnames within genealogy timeframes. SNPs (Single Nucleotide Polymorphisms) are stable, rare mutations used to define haplogroups (deep ancestry) and identify exact branches on a family tree.
Historically FTDNA has offered multiple STR-only Y panels at different marker counts; currently the main marketed levels are Y-37, Y-111, and Big Y-700 (which also includes STRs).
Estes, Roberta, STRs and SNPs – Are STR Markers Still Useful for Y DNA?, 3 Dec 2021, DNAeXplained – Genetic Genealology, https://dna-explained.com/2021/12/03/strs-and-snps-are-str-markers-still-useful-for-y-dna/
GeneaVlogger, DNA: SNP vs STR with Zach Gordon, 21 Aug 2017, Youtube , https://www.youtube.com/watch?v=hiooGmxzJAs&t=29s
A Comparison of Our Y-DNA Tests, FamilyTreeDNA Help Center, https://help.familytreedna.com/hc/en-us/articles/5579319716111-A-Comparison-of-Our-Y-DNA-Tests
Rowe-Schurwanz, Katy, What is Autosomal DNA? Beginner Guide to DNA Inheritance, 9 Apr 2026, FamilyTreeDNA Blog, https://blog.familytreedna.com/what-is-autosomal-dna/
[22] Mitochondrial DNA (mtDNA) tests analyze DNA inherited exclusively from the mother to trace direct maternal ancestry, identify deep ancestral origins, or diagnose genetic disorders. Because mtDNA changes very slowly and passes unchanged from mothers to all children, it is used for determining if individuals share a common maternal ancestor.
Mitochondrial DNA tests, This page was last edited on 11 October 2025, International Society of Genetic Gnealology Wiki, https://isogg.org/wiki/Mitochondrial_DNA_tests
[23] See for more information:
Connect and collaborate with genealogy enthusiasts. FamilyTreeDNA, https://www.familytreedna.com/group-project
Introduction to Group Projects, FamilyTreeDNA Help Center, https://help.familytreedna.com/hc/en-us/articles/4503173806351-Introduction-to-Group-Projects
Group Project Participation Informed Consent, 5 Jun 2018, FamilyTreeDNA, https://www.familytreedna.com/legal/terms/group-project-participation/06052018
Unkefer, Rachael, Four Types of Group Projects You Should Join, 23 Jul 2023, FamilyTreeDNA Blog, https://blog.familytreedna.com/group-project-categories/
Estes, Roberta, Project Groupings and How to Get the Most Out of Projects at Family Tree DNA, 21 May 2018, DNAeXplained – Genetic Genealology, https://dna-explained.com/2018/05/21/project-groupings-and-how-to-get-the-most-out-of-projects-at-family-tree-dna/
Estes, Roberta, FamilyTreeDNA Provides Y DNA Haplogroups from Family Finder Autosomal Tests, 30 Nov 2023, DNAeXplained – Genetic Genealogy, https://dna-explained.com/2023/11/30/familytreedna-provides-y-dna-haplogroups-from-family-finder-autosomal-tests/
Cloud, Janine, Which Group Projects Should You Join?, 18 Jan 2023, FamilyTreeDNA Blog, https://blog.familytreedna.com/group-project-types/
[24] Unkefer, Rachael, Four Types of Group Projects You Should Join, 10 Jul 2023, FamilyTreeDNA Blog, https://blog.familytreedna.com/group-project-categories/
Cloud, Janine, Which Group Projects Should You Join?, 18 Jan 2023, FamilyTreeDNA Blog, https://blog.familytreedna.com/group-project-types/
Introduction to Group Projects, FamilyTreeDNA Help Center, https://help.familytreedna.com/hc/en-us/articles/4503173806351-Introduction-to-Group-Projects
[25] The Group Time Tree: A New Big Y Tool for FamilyTreeDNA Group Projects, 15 Feb 2023, FamilyTreeDNA Blog, https://blog.familytreedna.com/group-time-tree/
[26] FTDNA describes these as geographic projects that “combine Y and mtDNA, and often Family Finder, for a comprehensive look at the genetic ancestry of a location,” which can range from an entire country down to a county, city, or shtetl.
ISOGG’s description, explicitly referencing FTDNA-hosted projects, defines a dual geographical DNA project as one that studies Y-DNA of men and mtDNA of both men and women from a specific location, sometimes with additional limits on surnames, heritage, or haplogroups; many such projects also collect autosomal data.
Core characteristics:
- Geographic focus: Membership is tied to ancestral roots in a defined place (e.g., a river basin, region, or country), not to one surname or single haplogroup, and can be as fine-grained as a small locality or as broad as a national project.
- Dual (or tri-) modality: By design, they integrate paternal lines (Y-DNA) and maternal lines (mtDNA); project descriptions often also invite Family Finder results to produce a more holistic picture of the region’s genetic structure.
- Examples in practice: ISOGG notes projects like the Alpine DNA Project and the New Zealand Dual Geographic Project at FTDNA, which accept Y-DNA, mtDNA, and atDNA, illustrating the model in live FTDNA projects.
Connect and collaborate with genealogy enthusiasts., FamilyTreeDNA, https://www.familytreedna.com/group-project
Geographical DNA projects, This page was last edited on 21 May 2024, Internaltional Society of Genetic Genealology Wiki, https://isogg.org/wiki/Geographical_DNA_projects
Dual geographical DNA project, This page was last edited on 31 December 2019,, International Society of Genetic Genealology Wiki, https://isogg.org/wiki/Dual_geographical_DNA_project
[27] Unkefer, Rachel, Four Type of Groups You Should Join, 10 Jul 2023, FamilyTreeDNA Blog, https://blog.familytreedna.com/group-project-categories/
Estes, Roberta, How to Join a Project at FamilyTreeDNA – And Why You Want To, 9 Nov 2021, DNA-eXplained – Genetic Genealogy, https://dna-explained.com/2021/11/09/how-to-join-a-project-at-familytreedna-and-why-you-want-to/
[28] See for example:
J. David Vance, J. David. “DNA Concepts for Genealogy: Y-DNA Testing Part 1.” YouTube, Oct 10 2019. https://www.youtube.com/watch?v=RqSN1A44lYU
Vance. “DNA Concepts for Genealogy: Y-DNA Testing Part 2.” YouTube, Oct 10 2019.
https://www.youtube.com/watch?v=mhBYXD7XufI
Vance. “DNA Concepts for Genealogy: Y-DNA Testing Part 3.” YouTube, Oct 10 2019.
https://www.youtube.com/watch?v=03hRXVg9i1k
Vance. “Automated from STRs, SNPs & Genealogies.” Genetic Genealogy Ireland 2017. YouTube. https://www.youtube.com/watch?v=2l8q2BJdTWI
Vance. “Vance/Vans/Wentz DNA Project Update October 2019.” YouTube, Oct 2019.
https://www.youtube.com/watch?v=5OIG_YHArB8
Vance. “The Case of the Clergyman’s Arms.” FamilyTreeDNA Blog, Jul 22 2024.
https://blog.familytreedna.com/case-clergymans-arms/
FamilyTreeDNA Blog. “Dave Vance Named FamilyTreeDNA General Manager.” Nov 27 2024.
https://blog.familytreedna.com/dave-vance-named-familytreedna-general-manager/
DNAeXplained (Roberta Estes). “Dave Vance Joins FamilyTreeDNA as Senior VP and General Manager.” Dec 3 2024.
https://dna-explained.com/2024/12/03/dave-vance-joins-familytreedna-as-senior-vp-and-general-manager/
Vance. The Genealogist’s Guide to Y-DNA Testing for Genetic Genealogy. 2020. Amazon/Kindle.
https://www.amazon.com/dp/B085HFBFD5
[29] Griffi(th)(n)(s)(ng), Background, FamilyTreeDNA, https://www.familytreedna.com/groups/griffith/about/background
[30] G-L497 Y-DNA, Background, FamilyTree, https://www.familytreedna.com/groups/g-ydna/about/background
[31] Welsh Patronymics, Background, FamilyTreeDNA, , https://www.familytreedna.com/groups/welsh-patronymics/about/background
[32] Wales Cymru DNA, Background, FamilyTreeDNA, https://www.familytreedna.com/groups/wales-dna/about
[33] 1 Feb 2022, email from Thomas Weaver to Jim Griffis, Subject: G-Z6748 and your Griffith/Griffis line
[34] 2 Feb 2022, email from Thomas Weaver to Jim Griffis, Subject: Re: G-Z6748 and your Griffith/Griffis line
[35] David Vance’s program is called SAPP – Still Another Phylogeny Program. It is a Y-DNA phylogeny builder that takes SNP and match data (especially Big Y) and infers a branching tree for a patrilineal cluster.
David Vance, The Life of Trees (Or: Still Another Phylogeny Program), https://www.jdvsite.com
[36] See: Rowlands, John and Sheila,The Use of Surnames Chapter Four, Patronymic Naming – A survey in Transition, Llanysul, Ceredigion: Gomer Press 2013
[37] As of the writing of this story, there were 153 FTDNA DNA testers that could trace their YDNA back to G-Z6748.
Your Haplogroup Story: G-Z6748, FamilyTreeDNA, Accessed 1 Apr 2026, https://discover.familytreedna.com/y-dna/G-Z6748/story













{kind=link}
{kind=link}